



Many people alive today still carry genetic traces of ancient human encounters. When modern humans left Africa and met Neanderthals and Denisovans, those meetings left a lasting mark in our DNA. Today, most people outside Africa carry around 2 percent Neanderthal ancestry, while some populations in Oceania also carry an additional Denisovan genetic legacy.
These inherited fragments are not just silent fossils inside the genome. They can influence skin biology, metabolism, immune reactions, disease risk, and how the body responds to infection. Some of these ancient variants may have helped early modern humans survive unfamiliar environments. Others may have become less useful as pathogens, lifestyles, and environments changed.
A new study now suggests that some inherited archaic DNA may be linked to weaker control of certain DNA viruses in modern humans. The research, published in Genome Biology and Evolution, examined whether genetic variants inherited from Neanderthals and Denisovans are associated with higher viral loads in people today.
The finding is especially interesting because earlier research often pointed in the opposite direction for RNA viruses. Several Neanderthal-derived immune variants have been linked to stronger responses against RNA viruses, which often cause acute infections and can spread rapidly. DNA viruses are different. Many of them can remain in the body for years, sometimes for life, often without causing obvious symptoms.
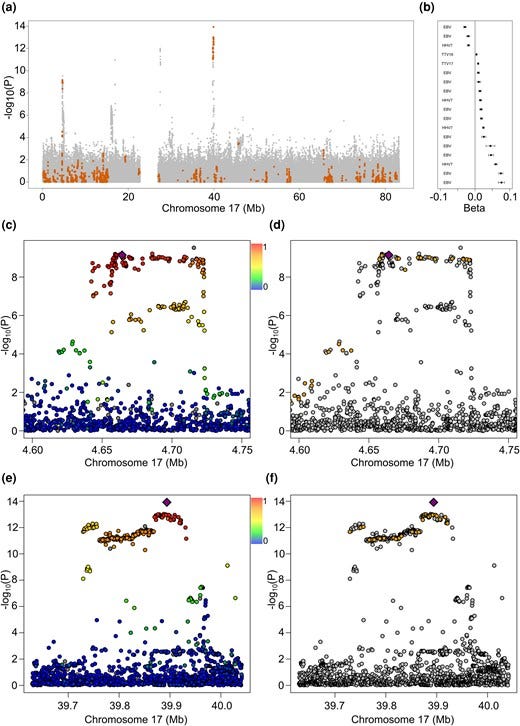
Figure 1. Archaic haplotypes associated with DNA virus levels. Panel a shows EBV level associations on chromosome 17, with Neanderthal marker variants highlighted in amber. Panel b presents effect size estimates from GWAS viral-load data for significant archaic SNPs, shown relative to the archaic allele and ordered by beta value. Panels c to f show two candidate regions on chromosome 17, centered on lead aSNPs chr17:4,664,269_T/C and chr17:39,893,872_C/T. These plots display linkage disequilibrium patterns in the UK Biobank data and highlight the distribution of archaic SNPs within each region.
That difference may be the key.
RNA viruses and DNA viruses do not behave in the same way. RNA viruses usually evolve quickly and are often connected with short, intense infections. DNA viruses tend to have more stable genomes and are more likely to persist quietly inside the body. They can hide in tissues, reactivate later, and interact with the immune system over long periods.
To explore this difference, researchers analyzed genome-wide association data from the UK Biobank. They focused on five common DNA viruses: Epstein-Barr virus, Human Herpesvirus 7, and three torque teno viruses known as TTV1, TTV16, and TTV17.
All five viruses are widespread in humans. Many people can carry them without visible illness. But even when symptoms are absent, viral load can still provide useful information. A higher viral load may suggest that the immune system is not controlling viral replication as effectively.
The team searched for links between these viral loads and archaic haplotypes, meaning groups of genetic variants inherited from extinct human relatives. These variants entered the modern human gene pool through interbreeding with Neanderthals and Denisovans tens of thousands of years ago.
The researchers identified 18 significant associations between archaic genetic regions and DNA virus load. These associations were spread across 13 distinct introgressed loci. Most were connected with Epstein-Barr virus, commonly known as EBV. Twelve of the associations involved EBV, four involved Human Herpesvirus 7, and two involved torque teno viruses.
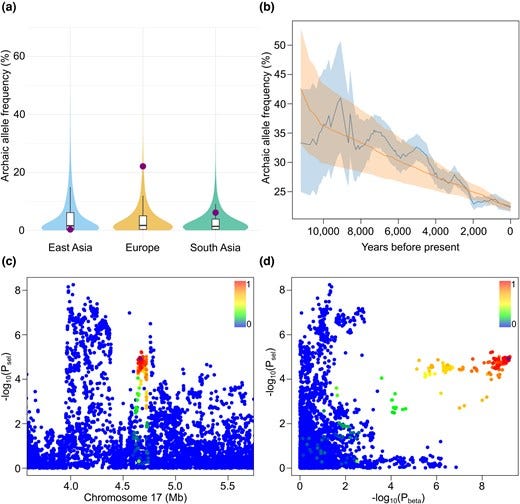
Figure 2. Evidence of selection linked to the archaic haplotype marked by aSNP chr17:4,664,269_T/C. Panel a shows the frequency of archaic haplotypes across three Eurasian populations, with the candidate haplotype highlighted in purple. Panel b traces the observed and modeled frequency of this archaic allele in ancient genomes from Europe and the Near East over the past approximately 11,500 years, including 95 percent confidence intervals. Panel c presents selection P-values for variants within a ±1 megabase region around chr17:4,664,269_T/C, colored by linkage disequilibrium and with the candidate variant marked by a purple diamond. Panel d compares selection signals and EBV association P-values across the same genomic region.
A striking pattern appeared: archaic variants were disproportionately associated with higher viral loads.
This does not mean that Neanderthal DNA directly causes disease. It also does not mean that everyone with these variants is at risk. The study points to statistical associations between inherited archaic DNA and the amount of viral DNA detected in large-scale genomic data. Still, the direction of the effect is notable, because it suggests that some ancient variants may not provide strong protection against certain persistent DNA viruses today.
Many of the associated regions were located in the Major Histocompatibility Complex, or MHC. This is one of the most important immune regions in the human genome. It helps the immune system recognize infected cells by presenting molecular fragments to immune cells. In simple terms, the MHC helps the body decide what belongs and what must be attacked.
Because the MHC is central to immune recognition, it is also one of the most complex and variable parts of the genome. This complexity makes interpretation difficult. Some archaic variants in the MHC may not be the direct drivers of the viral-load signal. Instead, they may be linked to nearby non-archaic variants with stronger effects. Even so, their presence in such an immune-critical region is biologically important.
One notable result involved a Denisovan-like haplotype associated with HLA-A*11:01, an immune-related allele that has previously been connected to EBV recognition by T cells. This archaic haplotype showed links with higher EBV load and appears at relatively high frequencies in East and South Asian populations. That pattern may point to a complicated evolutionary history, possibly including past positive selection.
The study also highlighted two Neanderthal-associated haplotypes on chromosome 17. Both were linked with higher EBV load. One of these regions showed possible functional connections with genes such as ARRB2, ALOX15, and PELP1. Another overlapped genes including GSDMB, IKZF3, ORMDL3, LRRC3C, and ZPBP2.
Several of these genes have immune relevance. GSDMB is involved in inflammatory programmed cell death, a process that can help clear infected cells. IKZF3 plays an important role in B cell biology, which matters because EBV infects B cells. ORMDL3 is connected with inflammation and cellular stress responses. These links do not prove a direct mechanism, but they show why these regions deserve closer study.
The evolutionary picture is not simple. A variant that appears disadvantageous today may not have been harmful in the past. Neanderthals lived in very different environments and faced different pathogen landscapes. The viruses circulating tens of thousands of years ago were not necessarily the same as those circulating today.
This means an ancient immune variant could have been helpful in one environment but less useful in another. A genetic tool shaped for one set of infections may perform poorly against a different set of viral challenges.
The study found evidence that one Neanderthal haplotype on chromosome 17 may have changed in frequency over time. In ancient West Eurasian genomes, the frequency of this archaic allele appears to have declined substantially over roughly the last 10,000 to 11,000 years. That decline may reflect more recent negative selection. In other words, the variant may once have been common or even useful, but later environmental and pathogen changes could have reduced its advantage.
This period overlaps with major changes in human life, including the spread of agriculture, denser settlements, closer contact with animals, and new patterns of infectious disease. Such shifts may have transformed the immune pressures acting on human populations.
The result is a more nuanced view of archaic DNA. Neanderthal inheritance is not simply good or bad. It depends on the trait, the environment, the pathogen, and the period in human history.
For RNA viruses, some inherited Neanderthal variants may have strengthened immune responses. For DNA viruses, especially persistent ones like EBV, some archaic variants may be linked to poorer viral control in modern humans. This contrast suggests that ancient admixture shaped immunity in different ways depending on the type of virus.
The study also has important limitations. Much of the available data came from people of European ancestry, meaning the analysis could not fully capture the diversity of Neanderthal and Denisovan DNA present in populations worldwide. Denisovan ancestry, in particular, is more common in some Oceanian and Asian populations, so broader datasets will be needed to understand the full picture.
The authors also emphasize that viral load is not the same as disease. A person can carry EBV or HHV-7 without symptoms. Higher viral load may reflect immune regulation, health status, inflammation, or other biological factors. More experimental work is needed to confirm whether these archaic variants directly influence viral control.
Still, the study adds an important piece to the story of human evolution. Our immune system was shaped not only by our own ancestors, but also by DNA inherited from extinct human relatives. Some of that inheritance may have helped modern humans adapt quickly when they entered new environments. Some of it may still influence how our bodies respond to infection today.
The legacy of Neanderthals and Denisovans is therefore not buried only in ancient bones. It remains active in living genomes, sometimes helping, sometimes complicating, and always reminding us that human biology is the result of a long and tangled evolutionary history.
Source article: Rajpara, R., Polishchuk, S. S., Yermakovich, D., & Dannemann, M. 2026. Archaic introgression shapes genetic variation at loci associated with DNA virus load in modern humans. Genome Biology and Evolution, evag110.